Abstract
Phylogenetic analysis of 16S rRNA gene sequences revealed that Enterococcus solitarius is not a member of the genus Enterococcus, but is related to species of the genus Tetragenococcus. On a phylogenetic tree, E. solitarius clustered with Tetragenococcus halophilus and Tetragenococcus muriaticus, with which it showed the highest 16S rRNA gene sequence similarity level (about 94 %). Phenotypic studies indicated that E. solitarius was also unable to produce acid from lactose, providing further evidence of its affiliation to the genus Tetragenococcus. DNA hybridization studies indicated that E. solitarius was clearly a separate species, different from T. halophilus and T. muriaticus (reassociation levels of about 23 and 54 %, respectively). As suggested in previous studies, E. solitarius is closely related to but clearly distinct from T. halophilus. Based upon properties that taxonomically distinguish it from species of the genus Enterococcus, it is proposed that E. solitarius be transferred to the genus Tetragenococcus and reclassified as Tetragenococcus solitarius comb. nov. (type strain, 885/78T=ATCC 49428T=CCUG 29293T=CIP 103330T=DSM 5634T=JCM 8736T=LMG 12890T=NCTC 12193T).
-
Published online ahead of print on 24 September 2004 as DOI 10.1099/ijs.0.63205-0.
-
The GenBank/EMBL/DDBJ accession number for the 16S rRNA gene sequence of Tetragenococcus solitarius DSM 5634T is AJ301840.
-
Results of DNA–DNA hybridization experiments are available as supplementary material in IJSEM Online.
Enterococcus solitarius was first described by Collins et al. (1989) and isolated from a human sample. The only strain known to date – the type strain – was identified on the basis of conventional physiological tests, as described by Facklam & Collins (1989). However, it was reported that strict adherence to these tests often led to misidentification of this particular species (Buschelman et al., 1993; Ruoff et al., 1990; Tsakris et al., 1998). With the use of 16S rRNA gene sequences in phylogenetic analyses, it was suggested that E. solitarius may in fact not be an enterococcal species, since its 16S rRNA gene sequence is too different from those of other Enterococcus species for it to be included in this genus (Collins et al., 1990; Patel et al., 1998; Williams et al., 1991). More recently, while performing sequence analysis of the 16S rRNA genes of lactic acid bacteria inhabiting paddy rice silage, we also noticed that E. solitarius was only remotely related to other enterococci (Ennahar et al., 2003). Other studies based on enterococci-specific PCR assays (Ke et al., 1999) or on whole-cell-protein profiles (Barros et al., 2001) were consistent with 16S rRNA gene sequence data. In fact, phylogenetically, E. solitarius appears to be more closely related to members of the genus Tetragenococcus (Barros et al., 2001; Collins et al., 1990; Ke et al., 1999; Patel et al., 1998; Williams et al., 1991), and it has even been suggested that E. solitarius and Tetragenococcus halophilus may constitute a single taxon (Ke et al., 1999; Monstein et al., 2001). However, controversy still exists, since a more recent study based on the sequence of a highly specific fragment within the enterococcal superoxide dismutase gene clearly placed E. solitarius among enterococcal species (Poyart et al., 2000). A comparative investigation including Enterococcus species and Tetragenococcus species has yet to be carried out to establish whether E. solitarius is a miscategorized taxon.
In this study, the E. solitarius type strain JCM 8736T was analysed with respect to its 16S rRNA gene sequence, DNA relatedness and phenotypic traits. In addition, T. halophilus JCM 5888T and Tetragenococcus muriaticus JCM 10006T, and the type strains of closely related Enterococcus species were included as references (Table 1⇓). Maintenance and propagation of cultures have been described previously (Ennahar et al., 2003).
Acid production from carbohydrates by E. solitarius, Tetragenococcus species and closely related species
Taxa: 1, Enterococcus casseliflavus JCM 8723T; 2, E. faecalis JCM 5803T; 3, E. gallinarum JCM 8728T; 4, E. cecorum JCM 8724T; 5, E. saccharolyticus JCM 8734T; 6, E. solitarius JCM 8736T; 7, T. halophilus JCM 5888T; 8, T. muriaticus JCM 10006T. All of the strains were positive for hydrolysis of aesculin and acid production from N-acetylglucosamine, d-fructose, d-glucose, d-mannose and salicin. None of the strains tested produced catalase or acid from adonitol, l-arabitol, erythritol, d-fucose, l-fucose, glycogen, 5-ketogluconate, d-lyxose, methyl xyloside, sorbose, l-xylose and xylitol. +, Positive; −, negative; (+), weak or delayed reaction.
E. solitarius JCM 8736T was compared with the type strains of Tetragenococcus species and closely related Enterococcus species using carbohydrate assimilation/fermentation on API 50 CH strips (bioMérieux), as described previously (Ennahar et al., 2003). Analyses of phenotypic features other than those analysed by the API system were performed as described by Cai et al. (1999). Our results indicate that, unlike other Enterococcus species, E. solitarius JCM 8736T did not ferment lactose, which is a common trait of both members of the genus Tetragenococcus (Table 1⇑). In addition, T. halophilus displayed the closest fermentation pattern to E. solitarius, with 18 identical reactions among the 28 differential carbohydrates tested. Both species produced acids from amygdalin, arbutin, cellobiose, galactose, gluconate, maltose, sucrose and trehalose, but not from glycerol, inulin, lactose, melibiose, methyl α-d-glucopyranoside, methyl α-d-mannopyranoside, d-raffinose, rhamnose, sorbitol or starch. The remaining carbohydrates yielded the same pattern of fermentation for all Enterococcus and Tetragenococcus species tested (Table 1⇑).
Phylogenetic analysis based on 16S rRNA gene sequences was carried out. Amplification, purification and sequencing were performed as described previously (Ennahar et al., 2003). The almost-complete 16S rRNA gene sequences were determined for E. solitarius JCM 8736T, T. halophilus JCM 5888T and T. muriaticus JCM 10006T. These were identical to sequences determined by other investigators (see GenBank/EMBL/DDBJ accession numbers in Fig. 1⇓). The highest levels of sequence similarity for E. solitarius were found with T. halophilus (93·8 %) and T. muriaticus (93·7 %), with values similar to that obtained between the two Tetragenococcus species (93·8 %). Similarity values between E. solitarius and other Enterococcus species were lower, generally ranging from about 88 to 90 %, while values between enterococci (except E. solitarius) ranged from about 95·0 to 99·1 %.
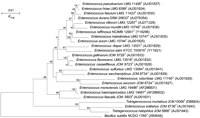
Phylogenetic tree showing the relative positions of Enterococcus species and Tetragenococcus species as inferred by the neighbour-joining method with 16S rRNA gene sequences. Bootstrap values for a total of 100 replicates are shown at the nodes of the tree. References of the type strains used for comparison are given, as well as the GenBank/EMBL/DDBJ accession numbers for all 16S rRNA gene sequences (in parentheses). Bacillus subtilis was used as an outgroup. Bar, 1 % sequence divergence.
Based on the 16S rRNA gene sequences, a phylogenetic tree was constructed by using the neighbour-joining method, as described previously (Ennahar et al., 2003). This analysis showed that E. solitarius formed a very well defined cluster with T. halophilus and T. muriaticus, with a bootstrap value of 100 % supporting monophyly (Fig. 1⇑). Among the two Tetragenococcus species, T. halophilus was the closest phylogenetic relative of E. solitarius in 94 % of bootstrap analyses, which reflects a higher 16S rRNA gene sequence similarity. The cluster making up the remaining enterococcal species was clearly distinct from the first one and was recovered in 98 % of bootstrap analyses. This finding, along with the phenotypic data, is in agreement with previous reports suggesting a close phylogenetic relationship between E. solitarius and Tetragenococcus species (Barros et al., 2001; Ke et al., 1999; Patel et al., 1998; Williams et al., 1991).
The relatedness of the genomic DNAs of E. solitarius JCM 8736T, T. halophilus JCM 5888T and T. muriaticus JCM 10006T was tested. Chromosomal DNA was purified by standard methods (Sambrook et al., 1989) as modified by Satomi et al. (1997). DNA–DNA hybridization experiments were performed by the method of Ezaki et al. (1989) using photobiotin labelling and colorimetric detection, as described previously (Ennahar et al., 2003). The results are presented in Table A, available as supplementary material in IJSEM Online. E. solitarius showed reassociation levels of 25·7 to 28·6 % with T. halophilus and 25·4 to 27·2 % with T. muriaticus. This is far lower than the DNA reassociation threshold value recommended for species delineation (70 %; Wayne et al., 1987). There were no great differences between the levels of homology measured when the genomic DNAs were used as targets or as probes. Therefore, as opposed to previous suggestions (Barros et al., 2001; Collins et al., 1990), E. solitarius constitutes a separate species, distinct from known Tetragenococcus species. On the other hand, DNA base composition, determined by HPLC as described previously (Ennahar & Cai, 2004), showed the G+C content for E. solitarius (38·3 mol%) to be only slightly different from those of T. halophilus (37·0 mol%) and T. muriaticus (36·5 mol%).
As described above, E. solitarius resembles Tetragenococcus species in its fermentative metabolism and 16S rRNA gene sequence. The low DNA–DNA reassociation values obtained for Tetragenococcus species clearly support the validity of E. solitarius as a separate Tetragenococcus species, with T. halophilus and T. muriaticus as its nearest phylogenetic relatives. Therefore, we conclude that E. solitarius (Collins et al., 1989) is a misclassified taxon that should be included in the genus Tetragenococcus as a separate species. The name Tetragenococcus solitarius comb. nov. is proposed.
Description of Tetragenococcus solitarius comb. nov.
Tetragenococcus solitarius (sol.i.tar.i′us. L. adj. solitarius alone, lonely).
Basonym: Enterococcus solitarius Collins et al. 1989.
Characteristics of the species are as described previously by Collins et al. (1989) with the following additions and corrections. Carbohydrate utilization pattern is as reported in Table 1⇑ of this study. The DNA base ratio is about 38 mol% G+C (as determined by HPLC). The GenBank/EMBL/DDBJ accession number for the 16S rRNA gene sequence is AJ301840.
The type strain is 885/78T (=ATCC 49428T=CCUG 29293T=CIP 103330T=DSM 5634T=JCM 8736T=LMG 12890T=NCTC 12193T).