Abstract
Until recently, there were only two recognised human diseases caused by Bartonella spp.: trench fever due to B. quintana and Carrion's disease due to B. bacilliformis. Since then, Bartonella spp. have been recognised as causative agents of further human diseases, including bacillary angiomatosis, cat-scratch disease, chronic bacteraemia, chronic lymphadenopathy, meningoencephalitis, stellar retinitis, myelitis, granulomatous hepatitis, endocarditis, osteomyelitis and peliosis hepatitis [1] (Table 1). In parallel, the genus Bartonella, which until 1993 contained only one species (B. bacilliformis), was broadly extended by reclassifying within it the genera Rochalimea [2] and Grahamella [3], and by the description of new Bartonella species (Fig. 1). Based on phylogenetic analysis of the 16S rRNA sequences, the relatedness of Bartonella spp. to other alpha-2 Proteobacteria including Brucella spp., Afipia spp., Agrobacterium tumefaciens, Bradyrhizobium spp. and Bosea spp. has been demonstrated [2, 3]. Active research on the pathogenesis of Bartonella infections has been triggered by the increased number of species of Bartonella, the re-emergence of older disease due to Bartonella, such as the modern form of trench fever that affects alcoholics and homeless people [4], the recognition of the role of Bartonella in AIDS-related diseases and the description of new clinical entities due to Bartonella spp. As a facultative intracellular bacterium, Bartonella interacts closely with its host cells. The study of these interactions gave an insight into some of the underlying virulence factors and pathogenic mechanisms, which may be common or specific to the host cell or to the Bartonella species studied, or both. Interactions of Bartonella spp. with (i) red blood cells and (ii) endothelial cells have been studied for several years, but (iii) bone marrow progenitor cells may also play a central role as a sanctuary site in the pathogenesis of Bartonella (Fig. 2).
- © 2002 Society for General Microbiology
Received 24 May 2002; accepted 4 June 2002.
Until recently, there were only two recognised human diseases caused by Bartonella spp.: trench fever due to B. quintana and Carrion's disease due to B. bacilliformis. Since then, Bartonella spp. have been recognised as causative agents of further human diseases, including bacillary angiomatosis, cat-scratch disease, chronic bacteraemia, chronic lymphadenopathy, meningoencephalitis, stellar retinitis, myelitis, granulomatous hepatitis, endocarditis, osteomyelitis and peliosis hepatitis [1] (Table 1). In parallel, the genus Bartonella, which until 1993 contained only one species (B. bacilliformis), was broadly extended by reclassifying within it the genera Rochalimea [2] and Grahamella [3], and by the description of new Bartonella species (Fig. 1). Based on phylogenetic analysis of the 16S rRNA sequences, the relatedness of Bartonella spp. to other alpha-2 Proteobacteria including Brucella spp., Afipia spp., Agrobacterium tumefaciens, Bradyrhizobium spp. and Bosea spp. has been demonstrated [2, 3]. Active research on the pathogenesis of Bartonella infections has been triggered by the increased number of species of Bartonella, the re-emergence of older disease due to Bartonella, such as the modern form of trench fever that affects alcoholics and homeless people [4], the recognition of the role of Bartonella in AIDS-related diseases and the description of new clinical entities due to Bartonella spp. As a facultative intracellular bacterium, Bartonella interacts closely with its host cells. The study of these interactions gave an insight into some of the underlying virulence factors and pathogenic mechanisms, which may be common or specific to the host cell or to the Bartonella species studied, or both. Interactions of Bartonella spp. with (i) red blood cells and (ii) endothelial cells have been studied for several years, but (iii) bone marrow progenitor cells may also play a central role as a sanctuary site in the pathogenesis of Bartonella (Fig. 2).
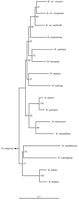
Fig. 1. Neighbour-joining tree resulting from comparison of sequences of groEL encoding genes of most Bartonella spp. identified to date. The values at each node represent the percentage of times each branch was found in 100 bootstrap replicates.
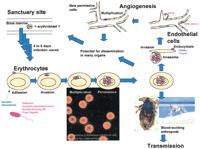
Fig. 2. Interactions of Bartonella spp. with red blood cells, endothelial cells and a putative sanctuary site located in the bone marrow. Bartonella and Bartonella-associated factors are represented in pink, host cell-associated factors are represented in green.
Bartonella infections in man and related clinical entities
The red blood cell
Infection of the red blood cell: a persistence and dissemination strategy
Bartonella infections are vector-borne diseases characterised by a natural cycle, vectors and reservoir hosts [5]. Human pathogenicity is either related to natural infection (B. quintana and B. bacilliformis) or to incidental infections (for other Bartonella spp.), acquired through contact with naturally infected mammals. Many Bartonella spp. have been shown to multiply and persist in red blood cells, sharing common persistence and dissemination strategies. Thus, B. henselae was found to infect cat red blood cells [6–8] and B. tribochorum those of rats [9], whereas B. quintana [10] and B. bacilliformis [11] were shown to invade human red blood cells. Bacteraemias were described even in healthy mammals, thus being one established exception to one of Koch's basic statements: ‘Bacteria do not occur in the blood or tissues of healthy animals or humans’ [12]. It was shown in the red blood cells of rats that the bacterial replication of B. tribocorum was regulated and stopped at a maximum of eight bacteria/cell [9]. The mechanism of regulation is unknown, but it prevents haemolysis and allows the persistence of Bartonella spp. within red blood cells. Disregulation of this mechanism, with uninterrupted multiplication of B. bacilliformis within red blood cells, might explain the rare acute haemolytic form of Carrion's disease (Oroya fever). However, as treatment of Bartonella with proteinase K has been shown to reduce the haemolytic activity of B. bacilliformis by 25%, a bacterial protein may also be involved in Oroya fever-associated haemolysis [13]. The ability of Bartonella to persist in erythrocytes for long periods may have been driven by evolutionary constraints, as it increases its transmissibility by blood-sucking arthropods. This persistence, which explains the prolonged bacteraemia in symptomatic and asymptomatic subjects, and the occurrence of disseminated disease in the homeless (chronic bacteraemia) or AIDS patients (bacillary angiomatosis), may also be favoured by the intra-erythrocytic localisation which partially protects Bartonella from the immune system.
Adherence to red blood cells
B. bacilliformis possess polar flagella (Fig. 3) that have been shown to mediate erythrocyte adhesion [14]. This flagella-associated adhesion was supported by the poor adherence of non-motile variants [11] and has been confirmed by the reduction of the erythrocyte-binding ability of B. bacilliformis by 40–50% with antiflagellin antibodies [15] and by the 75% reduction in binding ability of a flagellin-minus mutant [16]. Importantly, erythrocyte adhesion was not fully restored by complementation of the minus-mutant [16]. As the other Bartonella spp. (except B. clarridgeiae) do not possess polar flagella and as the B. bacilliformis flagellin-minus mutant retained some adhesive ability, other important mechanisms are involved in adherence to red blood cells. One of these may be the bundle-forming pili expressed at the surface of low-passaged B. henselae and B. bacilliformis cells, which have been implicated in the auto-agglutination propensity of Bartonella spp. [17]. Proteins exposed at the bacterial surface may also play a role in adhesion to erythrocytes, although no proteins able to interact with red blood cells have been identified yet. As adherence of B. bacilliformis to red blood cells is inhibited when the bacteria are pre-treated with N-ethyl maleimide, but is not inhibited when the erythrocytes are pre-treated with the same reagent [14], the red blood cell seems to play only a passive role in adherence. Membrane erythrocyte proteins, which bind B. bacilliformis passively, include spectrin and glycophorins A/B [18]. Enhanced binding of Bartonella to these proteins after treatment of erythrocytes with trypsin or neuraminidase has suggested a possible masking of binding site [18]. The fact that glycophorin A/B is the major receptor on human erythrocytes for Plasmodium falciparum merozoites [19] suggests that this protein is also utilised by Bartonella to bind to erythrocytes.

Fig. 3. B. bacilliformis and its polar flagella, within human erythrocytes, as seen by confocal microscopy. (Courtesy of J. M. Rolain, Marseille.)
Invasion of red blood cells
It has been speculated that the binding of Bartonella to spectrin may be a first step necessary to alter the erythrocyte membrane for internalisation of Bartonella cells [18]. This hypothesis is supported by the role of spectrin in the maintenance of erythrocyte shape and membrane integrity and deformability [18] and by the fact that proteases from P. falciparum were shown to cleave erythrocyte cytoskeletal components and spectrin [20]. A bacterial protein named deformin also appeared to be involved, at least for B. bacilliformis, in the formation of pits and trenches in the red cell membranes [11, 21], that may favour both colonisation and entry into the cell [17]. This molecule, initially thought to be a protein, seems to be a small hydrophobic molecule with affinity for albumin, although the effector mechanism remains to be elucidated [22]. The presence of a homologue of the deformin of B. bacilliformis that also led to the invagination of red blood cells, although less pronounced, has been identified in B. henselae culture supernate [23], suggesting that the deformin-mediated invasion mechanism might be shared between several Bartonella species.
At the molecular level, the genes of the invasion-associated locus (ial) identified in B. bacilliformis have been shown to confer an invasive phenotype to minimally invasive Escherichia coli strains [24]. The ial contains two genes named ialA and ialB. Immediately upstream lies a gene encoding a carboxy-terminal protease, ctpA, and another gene encoding the filament A polypeptide (FilA) [17, 25]. Downstream lie two open reading frames that do not share homology with other species [17, 25]. These six genes are thought to constitute a pathogenicity gene cluster, as (i) both ialA and ialB genes are needed to confer the invasive phenotype to E. coli; (ii) the gene encoding for the adhesion and invasion locus (Ail) protein of Yersinia enterocolitica, which is implicated in host cell attachment and invasion, shares 60% amino acid similarity with that encoded by ialB; and (iii) FilA exhibits amino acid similarity with various filamentous proteins including the M1 of Streptococcus pyogenes, involved in adhesion and invasion [17]. Furthermore, CtpA might also be involved in virulence, as a C-terminal protease of Salmonella typhimurium enhances intra-macrophage survival [26]. More importantly, several genes of this cluster have been sequenced in other Bartonella spp., and also in other facultatively intracellular bacteria, suggesting that some of the adhesion, invasion and persistence mechanisms used by Bartonella may be common to these species. Thus, the amino acid sequence of B. bacilliformis ialA shared 73% homology with that of B. clarridgeiae and 49% with that of Brucella melitensis, another member of the Rhizobium group, while the amino acid sequence of B. bacilliformis CtpA shared 81% homology with that of B. quintana and 71% with that of Br. melitensis (homology analysis performed with ClustalW) [27]. Whether ialA and ialB may explain the peculiar tropism of Bartonella for red blood cells remains to be defined. The role played by CtpA, FilA and both additional orphan open-reading frames remains also to be clarified. A role in quorum sensing, i.e., regulation of the intra-erythrocytic growth of Bartonella may be possible. In conclusion, it appears that specific (polar flagella) and common (deformin, invasion-associated locus) mechanisms have been developed by the different Bartonella spp. to adhere to and invade the red blood cells.
Angiogenesis
Apart from their tropism for red blood cells, a second typical pathogenic feature of Bartonella spp. is their ability to trigger angiogenesis. Such pathological angiogenesis is observed in bacillary angiomatosis and peliosis [28–31].
An unknown protein
The first experimental evidence that Bartonella-related angiogenesis may be due to a protein was afforded by Garcia et al. in 1990 [32]. They demonstrated that B. bacilliformis extracts possess an activity that stimulates endothelial cell proliferation up to three times that of a control [32]. The factor, which was found to be specific for endothelial cells and was larger than 12–14 kDa (not dialysed), was thought to be a protein because it was heat sensitive and precipitated with ammonium sulphate 45% [32]. B. bacilliformis extracts were also reported to stimulate the production of tissue plasminogen [32]. Live bacteria were later shown to increase both parameters (angiogenesis and tissue plasminogen production) in a fashion similar to the homogenates of B. bacilliformis [33]. In 1994, Conley demonstrated in a similar in-vitro model that B. henselae induces an angiogenic factor, whose susceptibility to trypsin also suggests that the factor may be a protein. More recently, it has been shown that this factor may be secreted by B. henselae [34], indicating that B. henselae may induce endothelial cell proliferation independently of bacterial invasion. The dramatic effect of erythromycin on the cutaneous lesions of bacillary angiomatosis [35, 36] may be due to inhibition of the production of the proteic angiogenic factor(s), as erythromycin is known to inhibit protein production at the ribosomal level. An angiogenesis-based effect would better explain the rapidity of its effect and its lack of a sustained response than a direct microbicidal one.
Vascular endothelial growth factor (VEGF)
VEGF, angiopoietins and ephrins were proved to be critical and specific for blood vessel formation [37]. Recently, Kempf et al. demonstrated that B. henselae induce EA.hy 926 cells (permanent endothelial cell line expressing factor VIII) to produce VEGF, which in turn was able to stimulate the proliferation of endothelial cells and the growth of B. henselae [38]. In the same study, Kempf et al. showed that the administration of VEGF-neutralising antibodies reduced endothelial proliferation by 50% [38], suggesting that other factors are involved, such as angiopoietins, ephrins or the yet unknown proteic factor discussed above.
Importantly, VEGF not only induces endothelial proliferation but also cell migration through several pathways that include the activation of a small GTP-ase RhoA, which is associated with phosphorylation of myosin light chain [39]. This may easily explain why Bartonella-associated angiogenesis is also characterised by altered spatial organisation within the monolayer and changes in cell morphology due to cytoskeleton re-organisation [40]. However, the impaired migratory ability of Bartonella-infected endothelial cells [41] was more surprising. This impaired mobility might result from the formation of thick robust stress fibres, probably via the activation of RhoA, as suggested by an inactivation assay [41]. Thus, the cells that effectively participate in angiogenesis are probably uninfected endothelial cells that respond to VEGF produced by the infected ones. B. henselae not only promotes the formation of new permissive cells but also protects the infected cells from apoptosis. Indeed, Bartonella suppress both early and late events in apoptosis, namely caspase activation and DNA fragmentation [42]. This anti-apoptotic effect, found to be specific for endothelial cells [42], may be mediated at least partially by VEGF, which is known to protect the cells from apoptosis [43].
Peliosis hepatitis
Peliosis hepatitis, which corresponds histologically to multiple blood-filled cystic spaces often communicating with the hepatic sinusoids, was initially thought to be induced by viruses [44]. Similar lesions were later observed in mice or rats exposed to various drugs or toxins, including phalloidine and oxazepam [45, 46] and reported in patients with advanced cancer or receiving anabolic steroids. The Bartonella-associated peliosis hepatitis, associated with HIV infection, differs from that of classical peliosis hepatitis by the additional presence of clumps of bacteria [47]. Recently, while studying the ability of VEGF to block tumour regression, Wong et al. observed that mice implanted with VEGF-expressing tumours sustained high morbidity and mortality that were out of proportion to the tumour burden. High serum levels of VEGF were associated with a lethal hepatic syndrome characterised by massive sinusoidal dilatation and endothelial cell proliferation and apoptosis [48]. A striking reversal of VEGF-induced liver pathology was achieved by surgical excision of VEGF-secreting tumours or by systemic administration of a potent VEGF antagonist [48]. As this VEGF-induced syndrome resembles cancer and Bartonella-induced peliosis hepatitis, the bacillary peliosis hepatitis may be directly due to the VEGF produced by the infected endothelial cells and by other infected cells that are known to secrete VEGF, such as erythroblasts [49].
Placenta growth factor
The placenta growth factor (PIGF) is analogous to VEGF and is secreted by the placenta and erythroblasts [37, 49]. Its effect on Bartonella multiplication remains to be defined, as does the role played by Bartonella in its production. However, the presence of vascular lesions in the maternal placenta of mice infected experimentally with B. birtlesii [50], and the role played by VEGF in Bartonella pathogenesis [38], suggest that PIGF might be involved in the genesis of the reproductive disorders observed by Boulouis et al. in infected mice.
Endothelial cells
The Bartonella–endothelial cell interaction is not restricted to angiogenesis stimulation. Thus, (i) invasion of endothelial cells was described for B. quintana [51], B. henselae [52] and B. bacilliformis [53], and (ii) a pro-inflammatory activation of endothelial cells was postulated, which is thought to be a result of receptor-ligand interactions between the activated endothelium and circulating neutrophils [54].
Adherence to endothelial cells
The bacterial ligands that may be involved in adherence may include bundle-forming pili (see above) and several outer-membrane proteins (OMPs). Of the nine proteins located in the outer membrane of B. henselae, five were shown to bind human umbilical vein endothelial cells [55]. Of these, a 43-kDa protein (Omp43), that exhibits a similar amino acid sequence to the Omp2b porin of Brucella spp., was shown to have especially high affinity with endothelial cells, suggesting that it may play a major role in Bartonella pathogenesis [55, 56]. The endothelial receptors involved in Bartonella adhesion may include intercellular adhesion molecule-1 (ICAM-1), which has been shown to be enriched especially at the tips of the protrusions of the endothelial membrane [52]. Interestingly, endothelial adhesion molecules (ICAM-1 and E-selectin) expression is upregulated via NF-κB translocation, induced by B. henselae [52].
Invasion of endothelial cells
Endothelial cells are invaded by two mechanisms: (i) endocytosis of bacteria, similar to that present in other intracellular clades, and (ii) the engulfment of clustered bacteria by a unique host cell structure called the invasome [52]. Invasion is associated with cytoskeletal re-arrangements, themselves induced by Bartonella via Rho GTPase signalling [41].
Type IV secretion system and the VirB operon
Type IV secretion systems consist of a multiprotein channel that transports DNA or protein from bacteria to host cell. Such a system is present in A. tumefaciens, another alpha-2 proteobacterium that parasitises plants. A gene cluster, the virB operon of A. tumefaciens, plays a critical role in the formation of the channel through which the transfer of oncogenic T-DNA occurs, resulting in tumour formation [58]. Type IV secretion systems coded by the virB operon are also present in Br. suis and Bordetella pertussis, where they appear to be required for survival in host macrophages and exportation of the pertussis toxin, respectively [59, 60]. Recently, Padmalayan et al. discovered that the gene encoding an immunogenic 17-kDa antigen of B. henselae was located within the virB operon of B. henselae [61]. The virB operon of B. henselae encodes 10 genes, of which eight share significant homology to those of A. tumefaciens, suggesting that it encodes for a type IV secretion system. Interestingly, no homologues of the gene encoding the immunogenic 17-kDa antigen are present outside Bartonella spp. [61]. Its expression is stimulated by endothelial cells [62]. Although the molecule that may be transferred to the host cell remains to be defined, some have speculated that it may be involved in the production of VEGF by the endothelial cells.
Activation of endothelial cells
A pro-inflammatory activation of endothelial cells was postulated, which is thought to be a result of receptor-ligand interactions between the activated endothelium and circulating neutrophils [54]. This is supported by the recent demonstration that B. henselae itself and B. henselae-derived OMPs induce an NF-κB-dependent upregulation of E-selectin and ICAM-1 in endothelial cells, which in turn results in enhanced polymorphonuclear rolling and adhesion [57].
Primary niche
In the rat model of B. tribochorum infection, the presence of periodic erythrocyte infection waves has been demonstrated [9], that echo the 5-day periodicity of the ‘Quintan fever'. The fact that Bartonella parasitise erythrocytes without leading to haemolysis, with the exception of B. bacilliformis, suggests that the re-infection waves are due to the liberation of the bacteria from a distant sanctuary site [54]. This unknown primary niche might be the endothelial cells, as suggested by Dehio [54]. However, a number of hints suggest that the primary niche or sanctuary site might instead be located in the bone marrow, as follows. (i) Although, in vitro, Bartonella is able to enter various cells, including macrophages [63], only erythrocytes and endothelial cells are permissive to Bartonella in vivo. The cells that play the role of primary niche should thus share some characteristics common to erythrocytes and endothelial cells. Candidates are mainly the cells that are issued by differentiation of the haemangioblast, the common precursor of both erythrocytes and endothelial cells. These include mainly angioblasts and erythroblasts. (ii) The fact that erythroblasts express VEGF [49], a factor known to enhance Bartonella replication [38]. (iii) All organs involved in bacillary angiomatosis could potentially play the role of sanctuary, including brain, penis, vulva, cervix, muscle and bone marrow [64–68]. However, after skin, bone is the second most frequent site, and bone lesions of bacillary angiomatosis are characterised by well circumscribed osteolysis, that is often painful and usually affects long bones [69]. Pain pattern and osteolysis echo those found in mastocytosis and multiple myeloma, suggesting that bone marrow cells may be infected by Bartonella. (iv) The outstanding features of trench fever are pain and tenderness in the shins and relapsing fever [70]. Many subjects also present only with painful shins, which were often, in the absence of fever, wrongly attributed to flat feet or rheumatism due to prolonged standing in mud and water [70]. This suggests that B. quintana may involve the bone marrow in trench fever patients. (v) The fact that bone marrow cells are highly permissive to Br. melitensis, a phylogenetically close relative of Bartonella spp. and to many other intracellular pathogens.
Conclusions
Future research should especially be aimed at defining the factors (i) determining the host specificity (especially for human-specific species), (ii) involved in erythrocyte and endothelial cell infection, and (iii) controlling Bartonella-associated angiogenesis. Moreover, the location in the host of a Bartonella sanctuary, if any, which could be responsible for the observed relapses of intra-erythrocytic infections should be identified. Bone marrow progenitors such as erythroblasts might be a potential candidate but this should be investigated further.